


Information über unsere Fossilien
© Henskens Fossils & John v. Straaten
PLEISTOZÄN
Mammuts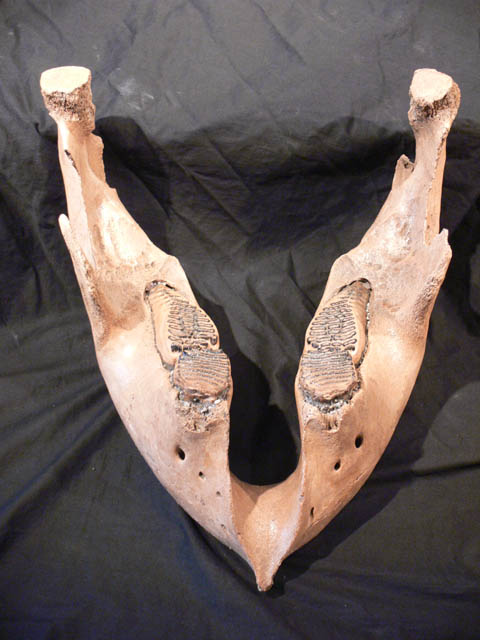
Vor ca. 55 Millionen Jahren tauchte eine Gruppe von Säugetieren auf,die wir unter dem Namen Rüsseltiere (Proboscidea) zusammenfassen. Rund 50Millionen Jahre später gingen aus dieser Abstammungslinie die ersten Mammutshervor. Die ältesten Funde von Mammuts sind etwa 4 Millionen Jahre alt undstammen aus Äthiopien. Sie gehören der Art Mammuthussubplanifrons an. Die Art überlebte sicher bis vor etwa 3 Millionen Jahrenund kurz darauf tauchte sein vermutlicher direkter Nachfolger M. africanavus in Nordafrika auf. DieseArt war wahrscheinlich der Vorfahre des Südelefanten (M. meridionalis).
Der Südelefant hatte mächtigeStoßzähne von beinahe 4 m lang. Sie zeigten schon die für Mammutscharakteristische Schraubenform. Große Exemplare des Südelefanten waren über 4m hoch. Er war das erste Mammut, das auch Eurasien erreichte. Vor etwa 1,5Millionen Jahren wanderte er über die damals trocken gefalleneBering-Landbrücke nach Nordamerika ein. Aus dem Südelefanten ging im UnterenPleistozän das Steppenmammut hervor.
Mit einer Schulterhöhe von bis zu4,7 m gehört das Steppenmammut (M.trogontherii) zu den größten bekannten Rüsseltieren. Es wird vermutet, dassdiese Tiere bis zu 10 Tonnen Gewicht erreichen konnten. Die spiralförmigenStoßzähne konnten bei den Männchen über 5 m lang werden. Das Steppenmammut wardie erste Mammutart, die durch starke Behaarung an das Leben in kalten Regionenangepasst war. Vor etwa 250.000 Jahren trat das Wollhaarmammut, das sich inSibirien aus dem Steppenmammut entwickelte, in Europe in Erscheinung, und lösteseinen Vorläufer nach einer kurzen Übergangszeit ab.
Das Wollhaarmammut (M. primigenius)wanderte aus Sibirien nach Europa und über die Bering-Landbrücke nachNordamerika ein. Namensgebend für diese Art ist das grobe Außenfell, das imWinter aus bis zu 90 cm langen Haaren bestand. Dieses Fell bildete an Bauch undFlanke eine Schürze. Unter diesen Deckhaaren befand sich eine dichteUnterwolle. Eine fast 10 cm dicke Fettschicht sorgte für zusätzlicheWärmeisolierung. Das Sommerfell war kürzer und weniger dicht als dasWinterfell. Eisleichen zeigen oft eine rötliche, relativ helle Fellfarbe. Andas kalte Klima war es auch durch die sehr kleinen Ohren angepasst.
Wollhaarmammuts waren nicht soriesig wie oft vermutet wird. Ausgewachsene Bullen erreichten eine Höhe von 2,8bis 4 m, nicht viel mehr als bei heutigen Elefanten. Die Tiere waren aberdeutlich massiger als Elefanten; das Gewicht betrug bis zu 8000 kg. Einauffälliger Unterschied zu den heute noch lebenden Elefantenarten sind diestark gedrehten Stoßzähne, eine kuppelförmige Beule auf dem Schädel, und derstark abfallende Rücken. Die Stoßzähne erreichten eine Länge bis zu 4,2 m. ImDurchschnitt sind sie aber 2,5 m lang. Kennzeichnend ist auch die hohe Lamellendichteauf den Backenzähnen.
Das bevorzugte Habitat war die sog.Mammutsteppe, eine Landschaft, in der sich Steppen- und Tundrapflanzenvermischten.
Man geht davon aus, dassWollhaarmammuts, ähnlich wie heutige Elefantenarten, in matriarchalisch geprägtenGruppen zusammenlebten. Das älteste Muttertier leitete die Gruppe, dienormalerweise aus 2 bis 9 Tieren bestand. Bullen dagegen führten eineinzelgängerisches Leben und kamen nur während der Brunft mit Weibchenzusammen.
Lange Zeit wurde angenommen, dieletzten Wollmammuts seien bereits vor 12.000 Jahren ausgestorben. Neue Fundezeigen, dass es erst vor 10.000 Jahren vollständig aus Nordeuropa verschwundenist. Die letzten Angehörigen lebten bis vor 3700 Jahren auf der Wrangel-Insel(Sibirien), wo sie wahrscheinlich durch das kleinere Nahrungsangebot eineSchulterhöhe von nur 1,8 m erreichten. Ob eine übermäßige Bejagung dasAussterben verursacht hat, oder rasche Klimaänderungen zum Ende desPleistozäns, ist bis heute umstritten.
Waldelefant (Elephas antiquus)
Der Waldelefant war mit seinemklimatischen Anspruch vermutlich auf mindestens sub-mediterranes Klimaangewiesen. Er lebte in parkähnlichen Landschaften und Laubwäldern. Die meistenüberlieferten Knochen- und Stoßzahnfunde stammen aus den letzten beiden großenZwischeneiszeiten. Bei der klimatischen Abkühlung am Ende der Warmzeiten zog ersich nach Süden in das wärmere Mittelmeergebiet zurück. In den Eiszeiten nahmdas Wollhaarmammut seinen Platz in Mitteleuropa ein, dessen Rückzugsgebietwährend der Warmzeiten in Nordosteuropa lag.
Der Waldelefant war mit bis zu 4,2 mSchulterhöhe größer als das Wollhaarmammut. Er hatte wahrscheinlich kein oderein kurzes Fell. Seine geraden Stoßzähne waren bis zu 3 m lang und nur am Endeetwas gebogen.
Die letzten Funde stammen aus demsüdlichen Spanien. Mit dem Rückzug der Art in den Mittelmeerraum kam es hiernach dem Ende der letzten Eiszeit zur Bildung zahlreicher Zwergformen. Ausursprünglich mit dem Festland verbundenen Landmassen bildeten sich mit demAnsteigen des Meeresspiegels Inseln. Die so isolierten Populationen bildeten inFolge von Nahrungsknappheit Zwergformen aus, auf u. a. Malta, Sizilien undZypern, die bis in die Bronzezeit überlebten.
Höhlenbär (Ursus spelaeus)
Der Lebensraum des Höhlenbären warEuropa, von Nordspanien bis zum Ural. Seine Kopf-Rumpflänge betrug bis zu 3,5m, seine Schulterhöhe zirka 1,7 m. Er war somit deutlich größer als der heutigeBraunbär. Die Weibchen waren, wie bei heutigen Bärenarten, etwas kleiner alsdie Männchen.
Der Höhlenbär hatte kräftige Kiefer,deren Muskeln auf der Oberseite des Kopfes an einem Scheitelkamm ansetzten.Seine großflächigen Zähne weisen jedoch darauf hin, dass er sich alsAllesfresser vermutlich hauptsächlich von Pflanzen ernährte. Er war keintypischer Vertreter der Kaltzeiten, weil er in Folge seinerErnährungsgewohnheiten nur bis zur nördlichen Grenze laubtragender Bäumeverbreitet war und in einem Tundren- und Kaltsteppenbiotop keine ausreichendeNahrung gefunden hätte.
Trotz der massenhaften Ansammlungvon Knochen und Zähnen in Höhlen war der Höhlenbär kein Höhlenbewohner. DieTiere halten in den Höhlen lediglich ihre Winterruhe, so dass es auch beimgelegentlichen Tod eines Tieres pro Höhle im Laufe Zehntausender von Jahren zugroßen Ansammlungen von Knochen und Zähnen kommt. Da die Knochen derHöhlenbären gelegentlich bis zu 90 Prozent aller Knochen ausmachen, trägt eineReihe von Höhlen die Namen Bärenhöhle oder Bärenloch. In der Drachenhöhle beiMixnitz (Steiermark) wurden Knochen von zirka 3000 Individuen freigelegt.
Derzeit sind jüngsteRadiokohlenstoffdaten über dieser Bärenart zirka 27.800 Jahren alt. DiesePeriode fällt mit einer Abkühlung des Klimas innerhalb des Eiszeitalterszusammen. Als Ursache für das Aussterben werden also Klimaänderungen und eindaraus resultierender Wandel der Vegetation (der Lebensgrundlage der Bären)vermutet. Der Höhlenbär starb damit schon vor der eigentlichen Aussterbewelleam Ende des Pleistozäns aus.
Wollnashorn (Coelodontaantiquitatis)
Das Wollnashorn lebte wohl alsEinzelgänger und dürfte mit einer Kopf-Rumpflänge van 3,5 m, einen Gewicht von3 t und zwei mächtigen Hörnern (das vordere bis zu 1 m lang) eine schwer zujagende Beute gewesen sein. Das vordere Horn war jedoch nicht rund imQuerschnitt wie bei heutigen Nashörnern, sondern flach und an der Vorderkantemeistens stark abgeschliffen. Man vermutet, dass das Tier mit seinem vorderenHorn die Schneedecke weggeschoben hat, um an die darunter liegenden Pflanzen zugelangen.
Seine Schädelform, vor allem das langgezogene Hinterhaupt, und seinehochkronigen Backenzähne lassen darauf schließen, dass es sich vorwiegend vonGräsern ernährte. Darauf deutet auch das breite Maul hin. Erhalten wurden dieseWeichteile bei mumifizierten Tieren aus dem sibirischen Permafrostboden. Andiesen Eisleichen fanden sich auch Fellreste, was uns neben der Überlieferungdurch Höhlenmalereien bestätigt, dass das Tier ein dichtes Fell besaß um sichvor der eiszeitlichen Kälte zu schützen.
Am Ende der letzten Eiszeit starbdas Wollnashorn aus. Ob Klimaänderungen oder menschliche Jagd oder beides fürsein Aussterben verantwortlich waren, ist umstritten.
Riesenhirsch (Megalocerosgiganteus)
Das Geweih des Riesenhirsches, dasnur die männlichen Tiere trugen, übertraf an Größe die Geweihe aller heutigenHirsche. Es erreichte eine Spannweite von 3,6 m. Er konnte, wie alle heutigenHirsche, das Geweih jedes Jahr abwerfen und neu aufbauen.
In Europa starb der Riesenhirschbereits vor 11.500 Jahren aus und war danach wohl auf Sibirien beschränkt. InWestsibirien überlebte die Art offenbar bis ins frühe Holozän und starb erstvor ca. 7600 Jahren aus. Nordamerika hat Megalocerosnie erreicht.
Steppenbison (Bisonpriscus)
Als Vorläufer desUr-Bisons gilt ein Rind der Gattung Leptobos.Dieser eurasische Bison entwickelte sich auf seiner Ausbreitung von Nordindiennach Westen und in den asiatischen Steppen zum Steppenbison weiter. Erüberquerte im frühen bis mittleren Pleistozän die Bering-Landbrücke undbesiedelte die nördlichsten Teile Nordamerikas.
Der Steppenbison starb am Ende desPleistozäns aus, aber eine kleinere Art aus Asien erreichte rund denselbenZeitpunkt Europa (der Wisent) und Nordamerika (der Amerikanische Bison).Steppenbisons wurden mehr als 2 m hoch. Die Breite zwischen den beiden Hörnernwar 1 m.
Auerochse (Bosprimigenius)
Der Auerochseentwickelte sich vor ca. 2 Millionen Jahren in Asien, und erschien inMitteleuropa vor rund 250.000 Jahren. Er hatte wahrscheinlich denselbenVorfahren als der Steppenbison. Er lebte in offenen Wäldern von Gräsern, Laubund Eicheln.
Mit einer Kopf-Rumpflänge von über 3m, eine Schulterhöhe von 1,75 bis 1,88 m bei den Bullen und einem Gewicht vonbis zu einer Tonne war der Auerochse bis zur letzten Eiszeit eines dermächtigsten Landtiere Europas. Der Geschlechtsdimorphismus war starkausgeprägt, die Kühe hatten eine Schulterhöhe von etwa 1,5 m. Die Hörner wurdenbis zu 80 cm lang, waren in typischer Weise nach vorn geschwungen und hatteneine schwarze Spitze. Die Angaben beziehen sich jedoch in erster Linie auf denvoreiszeitlichen Auerochsen. Nach der Eiszeit nahm der Auerochse in seinerGröße deutlich ab. Vor ca. 8000 Jahren wurde er auch domestiziert,wahrscheinlich im Bereich des heutigen Klein-Asien. Während er imMittelmeerraum und in Asien schon vor ca. 2000 Jahren ausstarb, war derAuerochse in Mitteleuropa sehr viel länger beheimatet. Durch Jagd undHabitatzerstörung wurde er immer seltener. Das letzte Exemplar starb in 1627 imWald von Jaktorow, Polen.
Höhlenlöwe (Pantheraleo spelaea)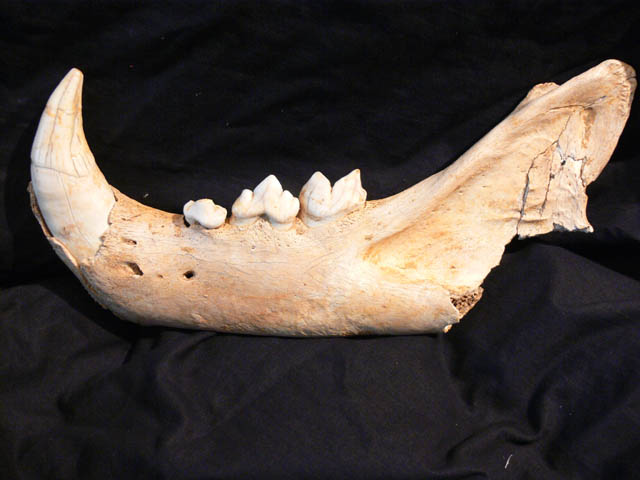
Der Löwe (Panthera leo) entwickelte sich vorungefähr 800.000 – 1 Millionen Jahren in Afrika. In Europa erschienen Löwenerstmalig mit der Unterart Panthera leofossilis. Mit einer Kopf-Rumpflänge von bis zu 2,4 m waren diese Löwen etwaum einen halben Meter länger als die heute in Afrika vorkommenden Löwen. Ausdieser riesigen Löwenrasse entwickelte sich schließlich der Höhlenlöwe, aucheine Unterart des Löwen, der zum ersten Mal vor etwa 300.000 Jahren auftrat.
Der Höhlenlöwe war weit über dasnördliche Eurasien verbreitet und drang selbst während der Kaltzeiten weit nachNorden vor. Über die durch die Vereisung trocken gefallene Bering-Landbrückeerreichte er auch Alaska. Von dort aus stieß er weiter nach Süden vor undentwickelte sich zum Amerikanischen Löwen (Pantheraleo atrox).
Europäische Höhlenlöwen waren etwa25 Prozent größer als heutige Löwen, erreichten aber nicht die enormen Ausmaßevon Panthera leo fossilis und Panthera leo atrox. SteinzeitlicheDarstellungen zeigen Höhlenlöwen immer ohne Mähne, was ein Hinweis darauf seinkann, dass männliche Tiere mähnenlos waren.
Löwen besiedelten Europa undNordasien sowohl in den Warmzeiten als auch in den Kaltzeiten. Ihre Nahrungbestand vor allem aus größeren Huftieren der damaligen Zeit, etwa Wildpferden,Hirschen, Wildrindern und Antilopen. Möglicherweise war der Höhlenlöwe ähnlichwie heutige Löwen ein Rudeltier.
Trotz seines Namens war derHöhlenlöwe kein dauerhafter Bewohner von Höhlen. Im Gegensatz zur Höhlenhyäneund zum Höhlenbär hat er Höhlen vermutlich nur selten als Versteck aufgesucht.Besonders kranke, alte oder geschwächte Höhlenlöwen suchten hier wahrscheinlichSchutz und verendeten.
Der Höhlenlöwe ist am Ende desPleistozäns ausgestorben. Bis dieser Zeitpunkt war der Löwe das weitverbreitetestegroße Landsäugetier neben den Mensch. Sie kamen vor in Afrika, der größte TeilEurasiens bis Indien, und in Amerika von Alaska bis Peru. Jetzt kommen sie nurin Afrika (südlich der Sahara) und in Nordwestindien vor.
Höhlenhyäne (Crocutacrocuta spelaea)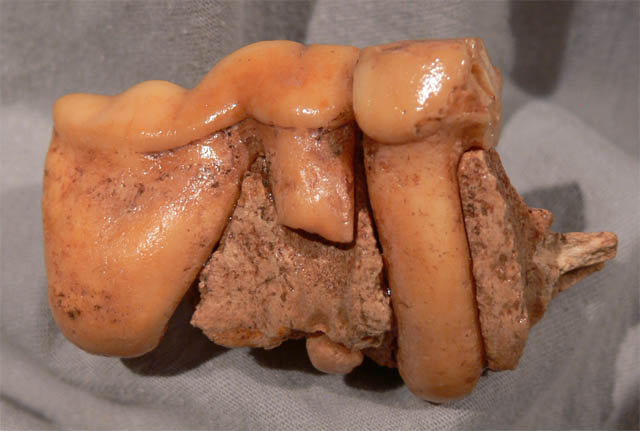
Die Höhlenhyäne war eine Unterartder Tüpfelhyäne (Crocuta crocuta),die während des späten Pleistozäns in Eurasien lebte. Sie nutzten Höhlen alsUnterschlupf, in dem sie die Knochen ihrer Beute sammelten. Die Höhlenhyäne waretwas größer als die heutige Tüpfelhyäne. Bei sowohl die heutige Tüpfelhyäneals die Höhlenhyäne sind die Weibchen etwas größer als die Männchen, was beiKarnivoren sehr ungewöhnlich ist.
Flusspferd (Hippopotamusmajor)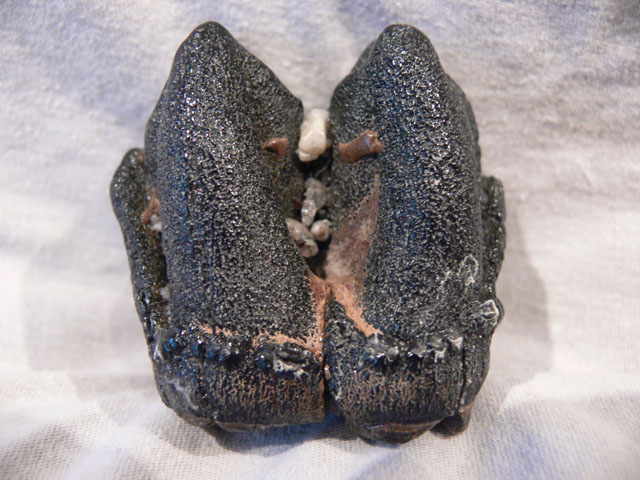
Die Flusspferde(Hippopotamidae) umfassen zwei lebende Arten, das (Groß-)Flusspferd und dasZwergflusspferd. Es sind schwerfällig gebaute, nahezu unbehaarte Säugetiere,die sich von Pflanzen ernähren und in Afrika südlich der Sahara leben.Traditionell werden sie in die Paarhufer eingeordnet, aber molekulargenetischeUntersuchungen und Fossilfunde haben zu der Sichtweise geführt, dass die Waledie nächsten lebenden Verwandten sind.
Die Flusspferde sind eineerdgeschichtlich junge Gruppe, die erstmals im Miozän vor etwa 15 MillionenJahren erscheint. Der älteste bekannte Vertreter ist Kenyapotamus aus dem mittleren Miozän aus Afrika. Im späten Miozänwar die Gruppe dann schon über Afrika und Eurasien verbreitet. Amerika habendie Flusspferde nie erreicht.
Bis ins Pleistozän kamen Flusspferdeauch in Europa vor, wobei sie nordwärts bis zum Rhein und nach Südengland vorstießen.Auf einigen Mittelmeerinseln entwickelten sich Zwergformen, die Beispiele fürInselverzwergung darstellen. Diese sind das Zyprische, Kretische, Maltesischeund Sizilianische Zwergflusspferd.
Ei eines Elefantenvogels (Aepyornis titan), Oberes Pleistozän, Madagaskar 
Die Elefantenvögel sind eine ausgestorbeneFamilie der Laufvögel mit den zwei Gattungen Aepyornis und Mullerornis.Sie ist durch Fossilien und zahl-reiche subfossile Eifunde von der InselMadagaskar vor der Ostküste Afrikas bekannt. Aepyornithiden wiesen die typischecursioriale Anatomie eines Bo-denvogels auf: Die Beine waren lang und die Zehenkurz, während der Flug-apparat nahezu gänzlich zurückgebildet wurde. DieArmknochen sind bis auf den Oberarmknochen verloren, das Brustbein ist flachund ungekielt. Der Hals ist lang und trägt einen relativ gesehen kleinenSchädel. Aepyornis maximus war dergrößte Vertreter des Taxons und dürfte zwischen 2,7 und
Aepyornithiden warenauf Madagaskar endemisch. Madagaskar trenn-te sich bereits in der Kreidezeitvon Afrika, was einige auf einen Verlust der Flugfähigkeit schließen ließ, daes keine großen Raubtiere auf Madagaskar gibt.
Man stimmtweitestgehend darüber überein, daß das Aussterben des Elefantenvogels aufmenschliche Einflüsse zurückzuführen ist. Wahrschein-lich wurden die großenVögel und deren Eier gegessen. Die von den Urein-wohnern betriebene Brandrodungspielte womöglich eine zusätzliche Rolle, da sie große Flächen des Lebensraumsdes Elefantenvogels zerstörte. Des Weiteren ist es möglich, daß Krankheiten,übertragen durch eingeführtes Ge-flügel, eine Rolle spielten. Das Datum desAussterbens der Elefantenvögel ist nicht sicher belegt. Archäologische Beweisebelegen ein Fortleben bis min-destens zum Jahr 1000. Gelegentlich wird über einÜberleben bis in das 17. Jahrhundert spekuliert. Der erste GouverneurMadagaskars, Étienne de Flacourt, berichtete von einem großen Vogel, welcherdie Eier eines Straußes legte. Dieser Vogel suche sich die entlegenstenRegionen, um nicht von Menschen bedroht zu werden.
Oft wird angenommen,daß die Aepyornithidae der Ursprung der Le-genden um den Vogel Roch sind. DieBezeichnung Elefantenvogel rührt von Marco Polos Beschreibungen des Roch her,in welchen ein Adler-ähnlicher Vogel von der Größe eines Elefanten geschildertwurde. Eier wurden möglich-erweise irrtümlich als die eines riesigenGreifvogels interpretiert, was Aepyor-nismaximus diesen Namen einbrachte.
TERTIÄR
Carcharocles megalodon
Der Megalodon ist eine ausgestorbene Haiartaus der Familie der Makrelenhaie, der weltweit gelebt hat. Dieser größtebekannte Hai der Erdgeschichte lebte im Miozän und Pliozän. Megalodon erreichte durchschnittlicheine Länge von 12 bis 14 Metern, wobei die größten gefundenen Zähne auf einTier von knapp 18 Metern schließen lassen. Gewöhnlich sind es bei fossilenKnorpelfischen nur die Zähne, die überdauert haben. Es gibt von Megalodon aber auch vereinzelteWirbelfunde, sowie Bissspuren auf Walknochen. Es wird davon ausgegangen, dasssich der Megalodon von Walenernährte.
GrubeMessel
In der Nähe der heutigen OrtschaftMessel entstand der Messeler Ölschiefer vor rund 47 Millionen Jahren. EineForschungsbohrung ergab, dass sich im Explosionstrichter eines Vulkanausbruchesein bis zu 300 Meter tiefer See, ein Maar, bildete, das durch Sedimentationwieder aufgefüllt wurde. Der Ölschiefer wurde in einer Stärke von bis zu 150Metern abgelagert. Das lässt auf eine Zeitspanne von rund 1,5 Millionen Jahrenschließen, während der dieser See bestanden haben muss. Die sehr große Tiefe imVerhältnis zur kleinen Oberfläche erlaubte nur in dem oberen Wasserschichteneinen Wasseraustausch durch Konvektion, was in der Tiefe zu Sauerstoffmangelführte. In dem subtropischen bis tropischen See konservierten sich deshalb in tiefergelegenen Wasserschichten und im Schlamm des Sees tote Tiere und Pflanzen, dieauf den Grund sanken und im Laufe der Zeit versteinerten.
In der Grube Messel wurde von 1859bis 1970 bituminöser Tonstein, Eisenerz und Braunkohle abgebaut und derTonstein zur Gewinnung von Erdölprodukten verschwelt. Nach Einstellung desÖlschieferabbaus war geplant, die Grube mit Müll zu verfüllen. Wissenschaftlerund die Bevölkerung engagierten sich für den Erhalt der Grube. Seit 1995 istsie in die Liste des Weltkultur- und Naturerbes der Menschheit von der UNESCOeingetragen.
Der Erhaltungszustand der MesselerFossilien ist herausragend. Es gibt allerdings ein Konservierungsproblem. DerTonstein enthält etwa 40 Prozent Wasser. Trocknet er aus, dann zerreißt er undzerkrümelt in kleine Blättchen. Erst seit Anfang der 1960-er ist es möglich,die Fossilien auf Kunstharz umzubetten und damit dauerhaft zu konservieren.
Green River Formation
Die Green RiverFormation ist ein Gesteinsverband aus dem Eozän in Nordamerika. Sie ist nachdem Green River benannt, einem Nebenfluss des Colorado Rivers. Sie besteht ausden Ablagerungen ursprünglich miteinander in Verbindung stehender großflächigerGebirgsseen. Im fossilträchtigen Gebiet um den sog. „Fossil Lake“ im SüdwestenWyomings treten zwei sehr feinkörnige Tonsteinlagen in Erscheinung, die eineVielfalt an vollständig und detailliert erhaltenen Fossilien enthalten.
Die Green-River-Fossilien sindungefähr 48 Millionen Jahre alt, wurden aber über einen Zeitraum von mehrerenMillionen Jahren hinweg abgelagert – dokumentiert wurde dabei der Übergang vomfeuchten Klima des frühen Eozäns zum etwas trockeneren mittleren Eozän. DasKlima war damals feucht und subtropisch warm, so dass selbst Krokodile, die keinenFrost vertragen, existieren konnten. Die Seen wurden von Platanenwäldernumrahmt, vereinzelte Palmwedel wurden entdeckt.
Die Lagerstätten bildeten sich unteranoxischen Bedingungen im feinkörnigen Kalkschlamm der Seen. MangelnderSauerstoff verlangsamte den bakteriellen Zerfall und hielt überdies Aasfresserfern, so dass Palm-, Farn- und Planatenblätter ungestört im feinkörnigenSediment eingebettet und erhalten werden konnten – einige Blätter zeigen sogarnoch die während ihres Wachstums erlittenen Insektenschäden. Insekten wurdenvollkommen erhalten, selbst Flügelmembranen sind noch zu erkennen.
Insgesamt wurden ungefähr 60Wirbeltiertaxa in der Green River Formation entdeckt. Neben den Fischen sind u.a. Reptilien, Vögel, Schildkröten und Fledermäuse vertreten.
KREIDE
Dinosaurier
Die Dinosaurierwaren die Gruppe der Landwirbeltiere, die von der Mittleren Trias bis zurKreide-Tertiär-Grenze die festländischen Ökosysteme dominierte. Die erstenDinosaurier spalteten sich vermutlich während der Mittleren Trias vor etwa 235Millionen Jahren von kleinen fleischfressenden Reptilien ab. Die Vertreteraller Dinosauriergruppen zeichnen sich durch eine Anzahl gemeinsamer Merkmaleaus, u. a. Schädelmerkmale und Merkmale des Schultergelenkpfannes und des Schienbeins.Das deutlichste Merkmal ist der Stand der Beine.
Bei den Dinosauriern standen dieBeine senkrecht unter dem Körper, ähnlich wie bei den meisten Säugetieren –aber anders als bei den meisten anderen Reptilien, deren Beine gespreizt nachaußen hin abstehen (Spreizgang). Durch ihre aufrechte Haltung konntenDinosaurier beim Bewegen leichter atmen, was wahrscheinlich Ausdauer- undAktivitätslevel erlaubte, welche die anderer Reptilien mit gespreizten Beinenübertrafen. Außerdem hat die gerade Stellung der Beine eventuell die Evolutiondes Gigantismus unterstützt, da so die Beine entlastet wurden.
Bislang wurden mehr als 500Gattungen und mehr als 9000 verschiedene Arten wissenschaftlich beschrieben. ImMittel kommen pro Jahr mindestens 30 neue Arten hinzu. Die Dinosaurier sindunterverteilt in Theropoden (zweibeinige Karnivoren), Sauropodomorphen (großePflanzenfresser mit langen Hälsen und Schwänzen), Ankylosaurier (vierbeinigePflanzenfresser mit massiven Hautpanzern), Stegosaurier (vierbeinige, Knochenplattentragende Pflanzenfresser), Ceratopsia (vierbeinige Pflanzenfresser mit Hörnernund Nackenschilden) und Ornithopoden (zwei- oder vierbeinige Pflanzenfresser).
Die Dinosaurier waren durch eineimmense Formenvielfalt gekennzeichnet. Einige waren Herbivoren(Pflanzenfresser), andere Karnivoren (Fleischfresser); einige waren vierbeinig,andere zweibeinig, oder beides. Viele hatten eine Panzerung, Hörner,Knochenplatten, Schilde oder Rückensegel. Obwohl sie für eine gigantische Größebekannt sind, variierte ihre Größe beträchtlich; so waren viele Dinosaurier nurso groß wie ein Mensch oder kleiner.
Das Wissen über Dinosaurier erhaltenPaläontologen durch die Untersuchung von Fossilien, die in Form vonversteinerten Knochen, Haut- und Gewebeabdrücken sowie durch Spurenfossilienüberliefert sind – also durch Fußspuren, Eier, Nester, Magensteine oderversteinerten Kot. Überreste von Dinosauriern sind auf allen Kontinentengefunden worden, einschließlich Antarktikas, da die Dinosaurier zu einer Zeitentstanden, als alles Festland im Superkontinent Pangaea vereinigt war.
Viele Dinosaurier (vielleicht alle)waren wahrscheinlich warmblütig. Der größte und schwerste Dinosaurier, derdurch gute Skelettfunde bekannt ist, ist der Sauropode Brachiosaurus. Ein Skelett, das aus den Knochen verschiedener etwagleichgroßer Individuen besteht, ist im Berliner Naturkundemuseum ausgestelltund hat eine Höhe von 12 Metern und eine Länge von 22,5 Meter; ein solches Tierhätte wahrscheinlich ein Gewicht von 30 bis 60 Tonnen gehabt. Der längste durchvollständige Skelette bekannte Dinosaurier ist Diplodocus, ein Skelettfund zeigt eine Länge von 27 Metern. Nochgrößere Sauropoden sind nur durch Skelettfragmente bekannt. Einer der größtenDinosaurier könnte Argentinosaurusgewesen sein, der auf ein Gewicht von bis zu 100 Tonnen geschätzt wird.
Unter den fleischfressendenDinosauriern gab es ebenfalls Riesen. Der größte durch fast vollständigeSkelettfunde bekannte Theropode ist der etwa 12 m lange Tyrannosaurus rex, jedoch gibt es hier ebenso Skelettfragmente, dieauf noch größere Gattungen schließen lassen. Der größte bekannte Theropode warvielleicht Spinosaurus mit einerLänge von 16 bis 18 m und einem Gewicht von 8 Tonnen.
Die kleinsten Dinosaurier hatten dieGröße eines Huhns. Die kleineren Dinosaurier ernährten sich fast ausschließlichkarnivor.
Die ersten Vögel tauchten im Juraauf und entwickelten sich wahrscheinlich aus den Maniraptoren, einer Gruppesehr vogelähnlicher Theropoden. Bereits 1861 wurde der berühmte Urvogel Archaeopteryx in den SolnhofenerPlattenkalken entdeckt, einer einzigartigen Fossillagerstätte in derFränkischen Alb. Dieser frühe Vogel weist ein Mosaik aus Merkmalen der Vögelund der Theropoden auf, ähnelt jedoch so sehr den Theropoden, dass mindestensein Fossil ohne klare Federabdrücken fälschlicherweise dem Compsognathus, einem kleinen Dinosaurier, zugeschrieben wurde. Bisheute wurden über 100 anatomische Gemeinsamkeiten zwischen theropodenDinosaurier und Vögeln nachgewiesen. Ähnlichkeiten zeigen sich besonders beimHals, Schambein, Handgelenk, Schultergürtel, Gabelbein und Brustbein.
Seit den 1990ern wurde eine Reihevon gefiederten Theropoden entdeckt, welche die enge Verwandtschaft zwischenDinosauriern und Vögeln noch deutlicher erscheinen lassen. Federn entwickeltensich ursprünglich wahrscheinlich zur Wärmeisolierung oder zur Zurschaustellung;die Funktion des Fliegens übernahmen sie erst später in der Evolution. Wie sichder Vogelflug entwickelt hat, ist jedoch noch nicht geklärt.
Am Ende der Kreidezeit kam es zueinem weltweiten Massenaussterben, das fast alle Tier- und vielePflanzengruppen erfasste, so auch die Dinosaurier. Zu den Ursachen gibt es verschiedeneVorstellungen. Die bekannteste Theorie ist ein Meteoriteneinschlag in Yucatan,Mexiko. Aber auch der enorme Vulkanismus bei der Entstehung derDekkam-Plateaubasalte in Indien könnte eine entscheidende Rolle gespielt haben.
Mosasaurier
Die Mosasaurierwaren in der Oberkreide eine weltweit verbreitete Gruppe meeresbewohnender Reptilien.Der erste Fund (von einem Schädel) war 1770 in einem Kalksteinbau beiMaastricht, Niederlande (daher kommt auch der Name Mosasaurier, was „Maasechse“bedeutet). Durch Truppen während der französischen Revolution kam das Fundstücknach Paris, wo der berühmte Naturforscher George Cuvier es als Echseidentifizierte. Er erkannte durch dieses Fossil erstmals in derWissenschaftsgeschichte, dass die Möglichkeit des Aussterbens von Lebewesenentsteht.
Die Mosasaurier stehenwahrscheinlich den rezenten Waranen sehr nahe und waren ein erfolgreicher, dochverhältnismäßig kurzlebiger Seitenzweig der Stammlinie der Warane, welcher sichder marinen Lebensweise anpasste. Sie erreichten bis zu 18 m Länge. Sieernährten sich wahrscheinlich von Fischen, andere Meeresreptilien undAmmoniten. Durch ein Gelenk im Unterkiefer konnte der Kiefer sehr weit geöffnetwerden um große Nahrung aufzunehmen. Der Augapfel wurde durch einen Ring ausdünnen Knochenplatten geschützt. Funden von Mosasaurierembryos innerhalb derKörper von ausgewachsenen Tieren geben an, dass Mosasaurier, genau wie dieIchthyosaurier, lebendgebärend waren.
Santana Formation, Brazilien
Die SantanaFormation ist eine der bedeutendsten Fossillagerstätten der Welt. Sie liegt inder Region Santana im Osten Brasiliens im Araripe-Becken. Die Fossilien stammenaus der frühen Kreidezeit vor etwa 110 Millionen Jahren. Die ersten Fossilienwurden von den beiden bayerischen Forschern Johann Baptist von Spix und CarlFriedrich Philipp von Martius 1819 entdeckt als sie diese damals nochunerforschte Region erkundeten und dabei auf sehr gut erhaltenen Fossilien vonFischen stießen.
Die Santana Formation bildete sichwährend der Unterkreide vor etwa 90 bis 114 Millionen Jahren. Zu dieser Zeitkam es zum Auseinanderbrechen des Riesenkontinents Gondwana, und die beidenTeile Südamerika und Afrika entfernten sich voneinander. Einige Fossilien die ausder Santana Formation bekannt sind (etwa Mesosaurus,ein kleines Süßwasserreptil) wurden außerdem an der Ostküste des afrikanischenKontinents gefunden, wodurch sie als Beleg für die Gondwana-Theorie angeführtwerden.
Quastenflosser Die Quastenflosser oder Coelacanthiformes sind eine Gruppe der Knochenfische in der Klasse der Fleischflosser (Sarcopterygii). Der Name bezieht sich auf die quastenförmige Ausbildung der sechs Ruderflossen. Ihre nächsten Verwandten sind die Lungenfische und die Landwirbeltiere. Die ältesten Fossilien stammen aus dem Unterdevon vor etwa 410 Millionen Jahren, die jüngsten aus der Oberkreide. Daher hat man angenommen, die Quastenflosser hatten das Massenaussterben am Ende der Kreidezeit vor 65 Millionen Jahren nicht überstanden. Bis 1938 aber eine rezente Art (Latimeria chalumnae) im Indischen Ozean vor Südafrika gefunden wurde. Deshalb gilt der Coelacanth als das bekannteste Beispiel eines lebenden Fossils. 1998 wurde vor der indonesischen Insel Sulawesi eine zweite Art (Latimeria menadoensis) entdeckt. Beide Arten werden bis zu 2 Meter lang und 100 Kg schwer. Fossil sind etwa 70 Arten bekannt. Der Bau der Brust- und Bauchflossen ähnelt dem Bau der Gliedmaßen der Landwirbeltiere. Vermutlich haben frühe Quastenflosser-Arten ihre muskulösen Flossen zur Fortbewegung am Meeresboden, vielleicht auch an Land benutzt. Die fossilen Quastenflosser hatten ein blasenartiges Organ am Darm, das als Rudiment einer auch als Lunge nutzbaren Schwimmblase interpretiert wird. Die Rhipidistia, nahe Verwandte der Quastenflosser und Lungenfische, werden als Vorfahren der ersten Landwirbeltiere angesehen.
Confuciusornis sanctus
Confuciusornis ist eine Vogelgattung deren fossilen Reste in derchinesischen Provinz Liaoning in Sedimentgesteinen der unteren Kreidezeitentdeckt wurden. Die ersten fossilen Überreste wurden 1993 entdeckt. Das Fossilwurde 1995 von einer chinesischen Forschergruppe als Confuciusornis sanctus („heiliger Konfuzius-Vogel“) beschrieben.
Confuciusorniswar so groß wie eine Taube und hatte einige Merkmalen, die bereits bei denVorfahren der Vögel (den theropoden Dinosauriern) ausgebildet waren (wie z.B.zwei Hinterhauptsöffnungen), aber auch Merkmalen, die im Verlauf derVogelevolution neu hinzukamen (wie z.B. mit dem Brustbein verbundenen Rippen).
Gemeinsame Merkmale aller Mitgliederder Familie Confuciusornithidae (1999 wurde eine zweite Art, Confuciusornis dui, beschrieben) sind das Fehlen von Zähnen, die nahezu viereckigeAusprägung des Deltopectoralkamms am Oberarm, der V-förmige hintere Rand desBrustbeins und die Größenverhältnisse der Handkrallen.
Viele Exemplare von Confuciusornis zeigen Abdrücke einesPaares verlängerter Schwanzfedern. Diese Fossilien sind sehr wahrscheinlich vonMännchen. Damit läge das früheste Indiz für Sexualdimorphismus bei Vögeln vor.
Hyphalosaurus
Hyphalosaurus ist eine ausgestorbene Gattung aus der KlasseChoristodera, Süßwasserreptilien die lebten vom Jura, möglicherweise der spätenTrias, bis mindestens das frühe Miozän. Hyphalosaurus-Fossilienwerden gefunden in der Jehol-Gruppe der frühen Kreide der Yixian-Formation inChina. Die positive und negative Platte des Holotypus (anhand dessen die Artzum ersten Mal wissenschaftlich beschrieben wurde) wurde an zwei verschiedeneGruppen von Untersuchern gegeben, die es beide einen verschiedenen Namen gaben:Sinohydrosaurus und Hyphalosaurus. Es wurde schon balderkannt, dass es sich um dasselbe Tier handelte, und da der Name Hyphalosaurus am ersten publiziertwurde, ist das der offizielle Name.
Hyphalosauruswurde maximal 80 cm lang. Er lebte im Wasser, was aus seinem langen Nackenund Schwanz und seinen verkürzten Beinen hervorgeht. In 2007 wurde ein Exemplarmit zwei Schädel gefunden, das altbekannteste Beispiel von siamesischenZwillingen.
JURA
DieSolnhofener Plattenkalke
Der Solnhofener Stein ausdem Oberjura war im Mittelalter schon weit verbreit und beliebt. Die römischen Baumeisterverwendeten den Plattenkalk beim Bau von Mauern, als Bodenbelag und fürInschriftentafeln. Die Steinplatten wurden auch später in Kelheim verladen undauf der Donau als sog. „Kelheimer Platten“ flussabwärts bis in diesüdosteuropäischen Länder verschifft. Größere Bekanntheit und Verwendungerlangte Solnhofener Plattenkalk durch die Erfindung der Lithographie im Jahre 1796, wofür dieFeinkörnigkeit des Plattenkalks Voraussetzung war.
Erst als zu Beginn des 20.Jahrhunderts der Steindruck durch moderne Druckverfahren abgelöst wurde,gewannen die Solnhofener Plattenkalke wieder mehr Bedeutung in derBauwirtschaft, insbesondere als Bodenpflaster, Wandverkleidungen,Treppenstufen, Fenstersimse und als Steine für Gartenmauern.
Bei der Gewinnung der Plattenkalkesind immer auch Fossilien ans Tagelicht gekommen. Zweifellos waren sie schonden Kelten und Römern bekannt. Das gezielte Sammeln und Erforschen derSolnhofener Fossilien begann erst mit Beginn des 19. Jahrhunderts. Einebedeutende Rolle spielten auch manche privaten Sammlungen in dieser Gegend.Neben den berühmten und sehr seltenen Fossilien von dem Urvogel Archaeopteryx lieferten die SolnhofenerSchichten im Laufe der Jahrhunderte eine Fülle weiterer fossiler Pflanzen undTiere. Bis heute sind etwa 750 verschiedene Arten beschrieben worden.
Bezeichnend für den Fossilinhaltist, dass nicht nur marine Formen wie Ammoniten, Krebse, Fische undMeeresreptilien nachgewiesen werden konnten, sondern auch Landpflanzen,Insekten, Schildkröten und Krokodile des Süßwassers, Brückenechsen,Dinosaurier, Flugsaurier und eben die Urvögel. Dies hängt mit der besonderenpaläogeographischen Situation des Ablagerungsraumes zusammen. Das Gebiet derheutigen südlichen Frankenalb war während der jüngeren Jurazeit eine Lagune.Sie lag am Südrand eines Festlandes und war durch einen Korallenriff-Gürtelgegen das offene Meer abgeschirmt.
Die Solnhofener Plattenkalke sindüber ein Gebiet von etwa 80 x 30 km in der südlichen Frankenalb, Bayern,verbreitet. Sie repräsentiert maximal eine halbe Million Jahre. Ihr absolutesAlter beträgt 150 Millionen Jahre.
Der Meeresboden desAblagerungsraumes der Solnhofener Plattenkalke war in einzelne Wannenunterteilt. Diese waren getrennt durch Erhebungen, die von altenMikroben-Schwamm-Riffen gebildet wurden. In den Wannen hatte sich eine Salz-und Dichteschichtung entwickelt mit einer lebensfeindlichen (sehr salzigen,sauerstoffarmen) Bodenzone stagnierenden Wassers und einer wenigersalzhaltigen, gut durchlüfteten Oberflächenzone. Dieses Bodenmilieu ohneAasfresser und eine wahrscheinlich rasche Einbettung führten zu derausgezeichneten Erhaltung von artikulierten Skeletten, manchmal sogar vonWeichteilen, die den Ruf der Solnhofener Plattenkalke als Konservat-Lagerstättebegründet.
Pterosaurier
Solnhofen isteine der weltweit bekanntesten Fossilfundstellen von Pterosauriern(„Flugsaurier“). Mindestens 17 verschiedene Arten wurden hier gefunden. Dieersten Pterosaurier entwickelten sich vor ungefähr 230 Millionen Jahren aus denArchosauriern (die auch die Vorfahren der Dinosaurier und Krokodile sind). DieFlugsaurier waren die ersten fliegenden Wirbeltiere. Sie hatten eine leichteBauweise mit hohlen Knochen und traten von der Größe eines Spatzen bis zuSpannweiten von etwa 12 m auf. Aus haarähnlichen Abdrücken bei einigenPterosaurierfossilien geht hervor, dass sie wahrscheinlich warmblütig gewesensind.
Das auffälligste Merkmal derFlugsaurier sind die zu großen Schwingen umgestalteten Vorderbeine, die denTieren sowohl den Gleitflug als auch den aktiven Flug ermöglicht haben. Dabeispannte sich die Flughaut zwischen dem Körper der Tiere und dem Arm mit demstark verlängerten vierten Finger der Tiere, der dem kleinen Finger beimMenschen entspricht, auf. Die anderen drei Finger bildeten Krallen außerhalbder Flughaut, der erste Finger fehlte.
Die Pterosaurier warenwahrscheinlich durchweg Jäger und Fleischfresser. Dabei jagten die frühenLangschwanzflugsaurier sicher vor allem nach Fischen und Kalmaren, und lebtenan Meeresküsten oder anderen großen Gewässern. Ein Indiz dafür sind die sehrlangen und reusenartigen Zähne, die perfekt zum Packen und Halten von Fischengenutzt werden konnten. Die Tiere lebten wahrscheinlich in größeren Kolonienähnlich den heutigen Brutkolonien von Meeresvögeln zusammen. Spätere kleineFlugsaurier waren allerdings sehr wahrscheinlich auch Insektenjäger.
Bei den Pterosauriern werden zweigrundsätzlich unterschiedliche Typen unterschieden. Dabei handelt es sich aufder einen Seite um die Langschwanzflugsaurier (Rhamphorhynchoidea) die sichdurch einen langen Schwanz und relativ kurze Mittelhandknochen auszeichnen.Diese Flugsaurier waren die frühesten Formen und verschwanden im oberen Jura.Ihnen werden die Kurzschwanzflugsaurier (Pterodactyloidea) gegenüber gestellt,die keinen oder einen sehr kurzen Schwanz und lange Mittelhandknochen besaßen.Erste Pterodactyloidea erschienen im Mittleren Jura und starben zum Ende derKreidezeit aus.
Pfeilschwanzkrebse
Pfeilschwanzkrebsewurden in den Solnhofener Plattenkalk verhältnismäßig oft gefunden. Siegehöhren zu den Gliederfüßern (Arthropoda). Der Fossilbericht derPfeilschwanzkrebse (auf English „horseshoe crabs“, Hufeisenkrabben, wegen ihrerhufeisenförmigen Körper) reicht 400 Millionen Jahre zurück. Deshalb werden sieauch als „lebende Fossilien bezeichnet. Sie müssen gut an ihren Lebensraumangepasst sein, sonst wäre es kaum möglich, dass sie sich über viele MillionenJahre gehalten und in den letzten 150 Millionen Jahren nur wenig veränderthaben.
Die Pfeilschwanzkrebse oderSchwertschwänze sind heute noch in vier Arten bekannt, die an deramerikanischen Atlantikküste und in Südostasien vorkommen. Sie werden bis zu 60cm lang. Der mit einem Panzer versehene Vorderkörper besitzt beiderseits eineErhebung, auf der zwei komplexe Facettenaugen sitzen, außerdem besitzen dieTiere zwei Punktaugen am Vorderrand des Panzers. Auf der Unterseite befindetsich der im Vergleich zum Panzer relativ kleinen Körper mit fünf einfachgebauten Beinpaaren, bei denen die ersten vier paar Beine in einer Schereenden. Das letzte Laufbein endet stattdessen in zwei Spitzen und besitztrelativ endständig blattförmig gestaltete Anhänge, die Flabella. Diese könnenden Kiemenraum sowohl abdichten als auch mit Atemwasser durchfächern. DerHinterleib endet in einem spitzen, starren Schwanzstachel.
Pfeilschwanzkrebse leben an den flachenSandküsten tropischer Meere in Tiefen zwischen 10 und 40 m. Normalerweise lebenPfeilschwanzkrebse auf dem Boden, können aber auch schwimmen. Sie ernähren sichvon Muscheln und anderen Weichtieren, die sie im Boden finden.
Der Posidonienschiefer bei Holzmaden
DerPosidonienschiefer oder Schwäbischer Ölschiefer ist ein asphaltgrauer bistiefschwarzer bituminöser Tonstein aus dem Unterjura. Das Gestein ist für seineaußergewöhnlich gute Fossilerhaltung bekannt, die den Posidonienschiefer zueiner Konservatlagerstätte von Weltrang gemacht haben. Der Begriff„Posidonienschiefer“ ist irreführend, da das Gestein weder Fossilien vonPosidonien (eine Gattung ausgestorbener Muscheln) enthält, noch impetrographischen Sinne ein Schiefer ist. Das Gestein ist nach einer häufigdarin vorkommenden Muschel benannt, die früher den Namen Posidonia bronni trug, aber heute einen anderen Namen hat. DerGattungsname Posidonia wurde auf eineGattung des Karbons übertragen. Und die scheinbare Schieferung ist eine sehrfein laminierte Schichtung des Sediments.
Vor ca. 175 Millionen Jahrenerstreckte sich über ein weites Gebiet des heutigen Mittel- und Nordwesteuropadas sog. Jurameer. Es entstand bei der Trennung Amerikas von Europa und Afrikaaus dem vormaligen Superkontinent Pangaea. Durch die eingeschwemmten Tone undKalke wurden die toten Tiere schnell überdeckt und konserviert. Im Laufe vonJahrmillionen verfestigte sich der Schlamm.
Das Posidonienschiefer ist bekanntfür die sehr gute Erhaltung von Skeletten. Am spektakulärsten sind dieIchthyosaurier erhalten, manchmal mit Hautabdruck oder Mageninhalt.Bemerkenswert sind außerdem die zahlreichen Weibchen, die während einesGeburtsvorganges oder mit Embryos im Bauch fossilisiert wurden. Das bezeugt, dassdie Ichthyosaurier im Gegensatz zu anderen Reptilien keine Eier am Land legten,sondern ihre Jungen, so wie sonst nur Säugetiere, lebend zur Welt brachten.Langhalsige Plesiosaurier und kurzhalsige Pliosaurier sind viel seltener. Etwaszahlreicher sind Krokodile. Von den Pterosauriern sind ungefähr 10 Exemplaregefunden. Bei den Fischen ist der Stachelhai Hybodus sehr bekannt.
Die Posidonienschieferplatten werdenzu Tisch- und Abdeckplatten, Wand- und Kaminverkleidungen, Treppenstufen undBodenplatten etc. verarbeitet. Das Gestein enthält wie die Ölschieferbituminöse Bestandteile und „schwitzt“ bei Wärme Schieferöl aus. Im 2.Weltkrieg wurde versucht aus Ölschiefer Treibstoff zu gewinnen.
Ichthyosaurier
Ichthyosauriererschienen in der Untertrias. Es waren Reptilien, die sich vollständig an dasLeben im Wasser angepaßt hatten. Sie lebten ausschließlich im Meer. Der Namebedeutet „Fischechse“. Sie sind sekundär wieder zum Leben im Wasser übergegangen.Sie ähnelten Delphine, hatten aber einen vertikalen statt horizontalen Schwanz,und vier statt zwei Schwimmflossen. Wie Delphine (Säugetiere die später auchnach dem Meer zürückkehrten) hatten Ichthyosaurier Lungen und mußten sie ab andzu an die Oberfläche kommen um zu atmen. Sie ernährten sich hauptsächlich vonFischen, Ammoniten und Belemniten. Insgesamt wurden etwa 80 Arten beschrieben. Dergrößte wurde
In dem Unteren Jura erreichten die Ichthyosaurier ihren größten Artenreichtum. Einebedeutende Fundstätte aus dieser Zeit ist der Posidonienschiefer bei Holzmaden.Bisher wurden die fossilen Überreste von etwa 3000 Exemplaren aus dreiverschiedenen Gattungen ausgegraben.Die Augen der Ichthyosaurier waren im Verhältnis zu ihrer Körperlänge sehrgroß, und der Stromlinienform des Körpers angepaßt. Wie die meisten Vögel undanderen Reptilien besitzen auch sie einen knöchernen Augenring (sgn. Sklerotikalring).Dieser Ring, der die Linse umfaßt, ist bei den Ichthyosauriern flach gewölbt,der äußere Rand ist nach innen umgebogen. Dadurch wird der rundliche Augapfelin eine nach außen abgeflachte Form gezwungen, wodurch das ungewöhnlich großeAuge kaum über die Körperoberfläche hervorragte.
Ichthyosaurier konnten nicht mehr an Land kriechen und dort Eier ablegen. Darumwaren sie lebendgebärend. Es wurden über 50 Fossilien gefunden, vor allem inHolzmaden, bei denen sich einige, maximal 11 Jungtiere im Körper desMuttertieres innerhalb des Rippenkorbes befinden.Die Ichthyosaurier starben vor etwa 90 Millionen Jahren aus. Die letzteGattung, die sich eindeutig identifizieren läßt, ist die weltweit verbreitete Platypterygius.Die Gründe für das Aussterben sind unbekannt.
 Stenopterygiussp., “DerSchlammtaucher“, Unterer Jura, Posidonienschiefer, Holzmaden, Deutschland
Stenopterygiussp., “DerSchlammtaucher“, Unterer Jura, Posidonienschiefer, Holzmaden, Deutschland
Da das Skelett weitgehend vollständig ist, hat dieserIchthyosaurier nach seinem Tode wahrscheinlich nur sehr kurz an derMeeresoberfläche gedriftet, bevor es agbesunken ist. Mit zunehmender Absinktiefewurden Brustkorb and Lungen immer stärker zusammengedrückt. Es erfolgteallmählich eine Verlagerung des Körperschwerpunktes Richting Kopf. Als Resultatdessen sanken solche Kadaver offenbar häufig kopfvoran dem Meeresgrundentgegen, mit einer Absinkgeschwindigkeit von ± 1,5 m/s. Bei dem Aufprall mitdem Meeresboden ist die Schnauze zersplittert, und wurde der Schädel stark inMitleidenschaft gezogen. Die Wirbelsäule ist ungefähr
Dieser Ichthyosaurier wird erwähnt in einem deutschenwissenschaftlichen Artikel über kopfwärtig gelandeten Ichthyosaurier (Der Ichthyosaur vomHauensteiner Nebelmeer, H. Hänggi, 2007, Naturforschende Gesellschaft desKantons Solothurn).
Ammoniten
Die Ammonitensind eine ausgestorbene Gruppe mariner Kopffüßer (Cephalopoda). Sie lebten vomUnterdevon bis zur Oberkreide. Die Gruppe war sehr artenreich; über 1500Gattungen sind bekannt, und über 30.000 verschiedene Arten. In der Mehrzahlliegt die Größe ausgewachsener Ammoniten zwischen 2 und 20 cm Durchmesser. Dengrößten bekannt gewordenen Riesenammonit stellt das Landesmuseum Münster aus.Es ist Parapuzosia seppenradensis,aus der Westfälischen Bucht. Sein Durchmesser beträgt 2,10 m. Ammonitenernährten sich wahrscheinlich von Plankton, Foraminiferen, Krebse,Stachelhäuter, Brachiopodenbrut oder Aas.
Das wichtigste und zugleich häufig das einzige erhaltene fossilierteMerkmal stellt die Schale der Tiere dar. Die Grundform des Gehäuses ist eine ineiner Ebene aufgerollte Spirale, wobei sich die Ränder der einzelnen Windungenmehr oder weniger umfassen. Die Schale ist in zwei Bereiche unterteilt, dieWohnkammer und den gekammerten Auftriebskörper. Die Kammern waren bei lebendenTieren mit Gas gefüllt. Der Regulation des Gases diente, wie bei den rezentenNautiloiden, ein Sipho, der allerdings anders als bei den Nautiloiden nicht inder Mitte der Trennwände, sondern an der Außenkante verläuft. Durch das Ein-bzw. Auslassen des Gases in die Septen war es dem Ammoniten möglich, im Wasserauf und ab zu steigen. Um ein Wachstum des Tieres zu ermöglichen, wurden zumeinen der Schalenrand an der Mündung vergrößert, und zum anderen im hinterenTeil der Wohnkammer neue Kammern abgetrennt.
Der Weichteilkörper befand sich hauptsächlich in der Wohnkammer und war beiNautilus mittels großer Muskeln mit ihr verwachsen. Außerhalb der Wohnkammerbefanden sich die Arme sowie der Kopf mit den Augen und dem Kiefer und dieMündung des Trichters. Die Ausbildung der Weichteile ist nur unvollständigbekannt, wahrscheinlich ähnelten sie im Aufbau denen der Nautiliden.
Ammoniten konnten sich im Wasser frei bewegen, allerdings bestimmt nichtsehr schnell. Grundlage der Bewegung war das Prinzip des Rückstoßes. Durch denTrichter wurde Wasser ausgestoßen. Dieser Rückstoß war wahrscheinlich nachhinten gerichtet, und der Kopf des Tieres zeigte somit in „Fahrtrichtung“.Normalerweise dürften Ammoniten über den Boden gekrochen sein oder sich knappdarüber bewegt haben.
Ammoniten fungieren als Leitfossilien, weil sie durch rasche Entwicklungimmer neue kurzlebige Formen hervorbrachten. Das bedeutet geringe vertikaleVerbreitung im Gestein und weltweite Verbreitung. Viele Gattungen und Artensind kosmopolitisch, haben weite Gebiete bewohnt und werden so in den einanderzeitlich entsprechenden Gesteinen der verschiedensten Länder gefunden.
TRIAS
Keichousaurus
Keichousaurus ist einSüßwasserreptil aus der Trias. Sein Name ist abgeleitet von derKweichow-Provinz (heute Guizhou-Provinz) in China wo das erste Exemplar 1958gefunden wurde. Keichousaurus war an ein Lebenim Wasser angepasst. Sie wurden maximal 30 cm lang, und hatten einen langenNacken und Schwanz und verlängerten Beinen mit fünf Fingern. Sie ernährten sichwahrscheinlich von kleinen Fischen, was aus den scharfen Zähnchen im Gebisshervorgeht. Sie gehören zu der Klasse der Sauropterygia („Eidechsen mitFlossen“), eine Gruppe erfolgreicher mariner Reptilien mit einer radikalenAnpassung des Schultergelenkes, die kräftige Schläge mit den Flossenermöglichte.
Nothosaurier sindmarine Reptilien aus der Gruppe der Sauropterygia („Flossenechsen“). Sie kamenwährend der mittleren und oberen Trias vor ums heutige Mittelmeer,Süddeutschland und in Südchina. Es waren Reptilien, die in Flüssen undküstennahen Gebieten lebten und sich auch an Land fortbewegen konnten. IhreVorderbeine hatten sich an die aquatische Lebensweise angepasst und wurdenstromlinienförmiger, die Hinterbeine sahen dagegen noch wie die Beine einesLandlebewesens aus. Der Schädel war flach und dreieckig, wie ein Krokodil. Diespitzen und gekrümmten Zähne im vorderen Bereich der Schnauze waren ziemlichgroß und die Zähne des Ober- und Unterkiefers griffen ineinander. Damit habendie Nothosaurier Fische gefangen. Die größte Art wurde fast 4 m lang.
Archegosaurus
Archegosaurus gehört zu den Labyrinthodontia, einer Gruppeausgestorbene Amphibien. Diese Amphibien waren die ersten Vertebraten die imDevon das Land eroberten. Sie haben sich aus den Sarcopterygii oderFleischflosser entwickelt, einer Klasse zu der die Quastenflosser undLungenfische gehören. In einer Gruppe der Sarcopterygii entwickelte sich inDevon die Schwimmblase zu Lungen als sie, aus unbekannten Gründen, dasMeereswasser verließen.
Archegosauruskonnte 1 m lang werden. Zusammen mit Sclerocephalusund Actinodon gehörte es zu dengrößten Amphibien in Europe während des Perms.
Discosauriscus
Discosauriscus ist eine Amphibie aus dem unteren Perm. SeineFossilien werden gefunden in Ablagerungen von Süßwasserseen in Mittel- undWesteuropa, vor allem in Tschechien. Die maximale Länge ist unbekannt, weil bisjetzt nur Fossilien von Jungtieren gefunden sind, nicht von erwachsenenExemplaren. Der Grund dafür ist wahrscheinlich, dass die erwachsenen Tierediese Seen benutzten um ihre Eier abzusetzen, aber selbst in einem anderenHabitat lebten. Es gibt zwei verschiedene Arten: D. austriacus und D.pulcherrimus. Von der letztere wurden nur einige Exemplare gefunden.
Sclerocephalus

Sclerocephalus war ein Salamander der im unteren Perm lebte indem Gebiet das heute Südwestdeutschland ist. Er lebte in Süßwasserseen und mitseiner Maximallänge von 2 Metern war er eine der größten Amphibien in Europa. Sclerocephalus ernährte sich von Fischenund anderen Amphibien. Genau wie andere Amphibien hatten die Larven von Sclerocephalus Kiemen um den Körper mitSauerstoff zu versorgen. Die erwachsenen Exemplare hatten Lungen und konntenalso zeitweilig aufs Land leben.
Micromelerpeton
Micromelerpeton war wie Sclerocephaluseine Amphibie der im unteren Perm lebte in dem Gebiet das heuteSüdwestdeutschland ist. Sie gehörte zu der Gruppe der Branchiosaurier:erwachsene Amphibien die wie eine Larve aussehen. Sie hatten z.B. externeKiemen und noch nicht verknöcherte Elemente im Hand- und Fußgelenk. Micromelerpeton konnte maximal 20 cmlang werden. Im unteren Perm lag Deutschland viel weiter südlich als heute, dieRegionaltemperatur war tropisch und das Klima war feucht; ideal für Amphibien.Die vornehmsten Feinde von Micromelerpetonin den permischen Süßwasserseen waren vor allem die größere Amphibie Sclerocephalus und Süßwasserhaie wie Orthacanthus.
Orthacanthus

Orthacanthus ist ein Süßwasserhai die im oberen Karbon undunteren Perm in Europa und Nordamerika lebte. Wegen seines langen Körpers ähneltes eine Muräne. Gerade hinter seinem Kopf hatte er einen Stachel. Orthacanthus konnte maximal 3 m langwerden. Er wurde 1889 zum ersten Mal beschrieben. Die weiße Farbe des Fossilsist ein Resultat der Erhitzung durch im Gestein aufsteigende Lava.
Er gehörte zur Familie derXenacanthidae, die erste Haie die auch das süße Wasser eroberten. Sie verlorenihre primitive Haiform und entwickelten einen aalartigen Körper. Fossilien vonkompletten Haien sind sehr selten da das Skelett aus Knorpel aufgebaut ist, wasviel weicher als Knochen ist.
KARBON
Farnpflanzen
Die Farne sindeine Gruppe von Gefäßsporenpflanzen, die die Schwestergruppe der Samenpflanzenbilden. Die Farne umfassen alle Gefäßsporenpflanzen außer den Bärlapppflanzen.Es gibt weltweit rund 12.000 verschiedene Arten. In Mitteleuropa sind etwa 200Arten bekannt.
Im Karbon bildeten Farne zusammenmit Schachtelhalmen und Bärlapppflanzen riesige Wälder und schufen somit dieBasis für die heutigen Steinkohle-Vorkommen. Die derzeitigen Farnpflanzenerreichten Baumgröße mit Höhen bis zu 30 m und fast 2 m Stammdurchmesser. Dieältesten fossilen Funde stammen aus dem unteren Devon. Sie sind somitwahrscheinlich älter als die Samenpflanzen, die erstmals im oberen Devonauftraten.
Farne sind weltweit verbreitet. Siekommen bis auf einige lichtliebende Arten fast ausschließlich an schattigen undfeuchten Plätzen vor. Den Schwerpunkt der Verbreitung haben sie in den Tropen.So findet man im tropischen Regenwald z.B. die größten Farnpflanzen, dieBaumfarne.
 Coccosteus cuspidatus
Coccosteus cuspidatus

Panzerfische
Als Panzerfischewerden mehrere Gruppen ausgestorbener fischartiger Wirbeltiere bezeichnet, dieim späten Silur und im Devon im Meer und in Süßgewässern lebten. Ihren Namenverdanken diese Tiere der starken Panzerung ihres Körpers mit hartem Gewebe.Der Begriff wird vor allem für zwei Gruppen verwendet: zum einen für dieOstracodermi („Knochenhäuter“), sowie für die Placodermi („Plattenhäuter“).
Als Ostracodermi werden urtümliche,kieferlose Fische bezeichnet. Sie kamen erstmals im oberen Silur vor, undstarben bereits im Devon wieder aus. Sie zeichneten sich durch einenKnochenpanzer am Kopf und einen Schuppenpanzer aus. Ihr Maul war klein, dieBrustflossen unbeweglich.
Die Placodermi sind eine Klassefischähnlicher kiefertragenden Wirbeltiere. Sie lebten vom oberen Silur an, undstarben am Ende des Devons wieder aus. Die frühen Placodermi warenSüßwasserbewohner, erst später besiedelten sie auch das Meer. Kopf und Rumpfwaren mit Knochenplatten gepanzert. Sie entwickelten als erste WirbeltiereKiefer. Wahrscheinlich entwickelten diese sich aus Knochen im Schlundraum, dievorher die Kiemenbögen der kieferlosen Fische stützten. Die größte Form war Dunkleosteus, der bis zu 6 m lang wurde.

Der Hunsrückschiefer bei Bundenbach
Weite Bereichedes Hunsrück und des Taunus im Gebiet von Mittelrhein und Mosel werden von sog.Hunsrückschiefern eingenommen, insgesamt ein Gebiet von mehreren 100 km². Essind dunkelgraue bis schwarze, siltige unter-devonische Tonsteine, die in über600 kleineren und größeren Dachschiefergruben seit dem Mittelalter abgebautwurden. Die Schieferplatten wurden von Hand gespalten und durchmustert. Hierbeiwurden marine Fossilien gefunden, die wegen ihrer Qualität insbesondere imGebiet von Bundenbach zu beliebten Sammelobjekten wurden. Eine ersteVeröffentlichung der Fossilien aus dem Hunsrückschiefer stammt von C.F. Roemer(1862 – 1864), der Asterozoen und Crinoiden der Sammlung des OberförstersTischbein beschrieb.
Aus der Tonschieferfazies desHunsrückschiefers wurden 6 Pflanzenarten und mehr als 260 Arten tierischerFossilien beschrieben. Unter den Tierarten befinden sich Stachelhäuter,Seelilien, Sterntiere, Trilobiten, und Fische (u. a. zwei Gattungen vonkieferlosen Panzerfischen). Insbesondere hat die Möglichkeit, Fossilien imRöntgenbild sichtbar zu machen, zur entsprechenden Untersuchung derHunsrückschiefer-Fossilien angeregt. Ein Pionier auf diesem Gebiet war W.M.Lehmann (1880 – 1959), ein Physiker, der als Honorarprofessor amPaläontologischen Institut der Universität Bonn arbeitete.
Sowohl um die stratigraphischeEinordnung und fazielle Interpretation der Hunsrückschiefer als auch um dieLebens- und Ablagerungsbedingungen im „Hunsrückschiefermeer“ gibt es Kontroversen.Die Deutungen reichen von einem mehrere 100 m tiefen, vollmarinenAblagerungsraum bis zu einem Wattenmeer-Modell, von der Annahme gewöhnlichlebensfeindlicher Bedingungen am Meeresboden über episodische Massenvergiftungder Organismen bis zu lebensfreundlichen Bedingungen in gut durchmischtem,reich belebtem Wasser.
Seeskorpione
Seeskorpione (Riesenskorpione,Eurypteriden) sind eine ausgestorbene Gruppe von Kieferklauenträgern. Sie warenWasserbewohner und lebten vom Ordovizium bis zum Perm. Ihre Fossilien werdenfast weltweit gefunden in Ablagerungen von sowohl Meeres-, Brack- alsSüßwasser. An Land entstandene Bewegungsspure deuten darauf hin, dassSeeskorpione das Wasser zumindest kurzfristig verlassen konnten. Mit über 2 mKörperlänge stellen einige Arten die größten bekannten Gliederfüßer überhauptdar.
Die Eurypteriden hatten einenschmalen Panzer mit einem nur wenig gegliederten Kopfschild und einemHinterleib mit zwölf Segmenten. Die Beine und anderen Gliedmaßen setzten wiebei den heutigen Spinnen und ihren Verwandten ausschließlich am Kopfschild an,das hintere Beinpaar war zu großen Schwimmbeinen umgebildet. Die vorderstenBeinglieder sowie die Cheliceren (Kieferklaue) waren mit zangenartigenStrukturen und Zähnen bestückt, die ein Festhalten der Beute ermöglichte.
SILUR
Graptolithen
Die Graptolithen(„Schriftsteine“) sind eine ausgestorbene Klasse polypenähnlicher,koloniebildender mariner Tiere. Fossil überliefert sind nur die Wohnröhren, dieeinen Kammeraufbau aufweisen. Die Lebensweise der Graptolithen war zu Beginnsessil-benthisch (im Meer lebend, festsitzend). Im weiteren Verlauf derEvolution entwickelten sich planktonische Arten. Die Bauweise der Graptolithenwies ein Wachstum entlang einer oder mehreren Achsen auf. Die Kolonien hatteneinen oder mehrere Achsen mit geraden oder gebogenen Wuchsformen. Aufgrund derzeitlich raschen Entwicklung der Klasse, ihrer weiten Verbreitung und denmakroskopisch leicht erkennbaren Fossilien sind die Graptolithen ausgezeichneteLeitfossilien vom Oberkambrium bis ins Unterdevon.
ORDOVIZIUM
Cystoidea
Die Cystoidea(Beutelstrahler) sind eine ausgestorbene Klasse innerhalb des Stammes derStachelhäuter und sind entfernt verwandt mit den noch heute lebenden Seelilienund Haarsternen. Es gibt etwa 100 bekannte Gattungen. Sie werden gefunden inEuropa, Nordamerika und Australien. Sie entstanden im oberen Kambrium. Diegrößte Formenfülle hatten sie im Ordovizium. Im Silur und Devon verlieren siemehr und mehr an Bedeutung, die letzte Art verschwindet im Oberdevon.
Typisch für die Beutelstrahler istein Porensystem, dass vermutlich der Atmung diente. Es war entweder paarigangeordnet oder aus Porenrauten aufgebaut. Die Beutelstrahler ernährten sich,indem sie fressbare Partikel mit ihren Brachiolen aus dem vorbeiströmendenWasser filterten und über Futterrinnen zum Mund führten.
Manche Beutelstrahler waren ständigmit einem Stiel am Meeresboden befestigt, andere haben ihren Stiel um einengeeigneten Gegenstand geschlungen und konnten vielleicht ihren Standortwechseln. Andere Formen waren ungestielt und hafteten am Untergrund odersteckten im Sediment.
KAMBRIUM
Trilobiten
Die Trilobiten(“Dreilapper”) sind eine ausgestorbene Klasse meeresbewohnender Gliederfüßer (Arthropoda). Der Name Trilobit bezieht sich auf die Dreiteilung des Panzers in Cephalon(Kopfschild), Thorax (Rumpf)und Pygidium (Schwanzschild) und quer dazu die außen liegenden Pleuren(Seitenteile) und die mittig liegende Achse (Rhachis, Spindel). Sie existiertenvom mittleren Kambrium bis zum Massenaussterben am Ende des Perms (520 – 250Millionen Jahren). Ihre mit Calciumcarbonat zu einem Panzer verstärktenExoskelette sind als Fossilien in großer Zahl erhalten geblieben.
Die Klasse der Trilobita besteht aus9 anerkannten Ordnungen, über 150 Familien, über 5000 Gattungen und mehr als15.000 beschriebenen Arten. Jedes Jahr noch werden weitere Arten gefunden undbeschrieben. Der kleinste bekannte Trilobit hat nur 1 mm Länge, der größte 70cm. Die Meere wurden vom strandnahen Flachwasser bis in Tiefen von ca. 300 mbewohnt. Die helleren, oberflächennahen Bereiche bevorzugten die Arten mitguten Sehvermögen, während jene mit schwachem oder ohne Sehvermögen wohl in dentiefen und lichtarmen Meeresteilen zu Hause waren oder eingegraben lebten.
Nicht alle Trilobitenarten habenAugen ausgebildet. Sind Augen vorhanden, so handelt es sich um Facettenaugen,die wie das Exoskelett aus Calcit bestehen. Die Spaltbeine waren zweiästig. Dererste Ast diente der schwimmenden Bewegung im Meer. Dazu hatte es fächerartigeErweiterungen. Der zweite Ast diente dem Gehen auf dem Meeresgrund.
Trilobiten hatten keinedifferenzierten Kauapparate. Sie waren also wohl nicht in der Lage, harte oderzähe Nahrung zu zerkleinern. Wir dürften deshalb annehmen, dass die Tiere vonPflanzenmulm, Kleintieren oder von Aas lebten.
Trilobiten zählen zu den wichtigstenLeitfossilien der Erdgeschichte, nicht nur weil sie fast weltweit vorkommen.Ihre Überreste werden zu relativen Altersbestimmung von Sedimentgesteinen genutzt.Diese Methodik nennt man Biostratigraphie. Bestimmte Trilobitenarten kommen nurin engen zeitlichen Abschnitten vor, und sind also für die Ablagerungen dieserZeit kennzeichnend.
Stromatolithen
Stromatolithensind fossile Algenmatten von Kalkgesteinen, die im Millimeterbereichgeschichtet sind und eine abwechselnde Hell-/Dunkel-Färbung aufweisen. DieHellfärbung ist auf magnesiumhaltigen Kalzit zurückzuführen, der in derumhüllenden Schleimschicht der Algen abgelagert ist. Die Dunkelfärbungresultiert aus herabrieselnden Sedimentpartikeln, die sich im Algengefechtverfangen. Sie werden weltweit gefunden.
Wesentliche Voraussetzung derBildung von Stromatolithen sind Biofilme, die aus Mikroorganismen bestehen(Mikrobenmatten). Die Bildung von Stromatolithen wird begünstigt oder sogarerst ermöglicht, wenn der für ihre Bildung erforderliche Biofilm nicht durchandere Organismen abgefressen wird. Bei sehr alten Stromatolithen desPräkambriums war das gegeben, weil es noch keine derartigen Lebewesen gab. Ausdem Präkambrium sind deshalb Stromatolithen zahlreich bekannt. Bei jüngerenfossilen und rezenten Stromatolithen war bzw. ist dies nur in Milieus der Fall,die für andere Lebewesen ungünstig sind, z.B. bei hohem Salzgehalt.
Stromatolithen werden als dieältesten Fossilien bezeichnet und kommen schon im Präkambrium vor, sieexistieren seit etwa 3,5 Milliarden Jahren. Sie werden als die ersten erkennbardurch Organismen aufgebauten Strukturen gesehen. In manchen Zeitabschnitten derErdgeschichte – lange bevor es Korallen gab – waren die Stromatolithenbildnerwichtige Riffbildner. Bis vor einer Milliarde Jahren waren Stromatolithen infast allen Küstengewässern stark verbreitet. Vor 700 Millionen Jahren ging dieVielfalt und die Verbreitung der Stromatolithen abrupt stark zurück. Sehrselten werden Stromatolithen ab 450 Millionen Jahren vor der Gegenwart. Mannimmt an, dass sie von den aufkommenden mehrzelligen Eukaryoten massenweiseabgeweidet und so verdrängt wurden.